|
Proszę chwilę zaczekać, ładuję stronę ... |
|
Proszę chwilę zaczekać, ładuję stronę ... |
|
||||||
|
5.2. Badania dynamiki zmian poziomu wód podziemnych oraz ich wpływu na podmokłe ekosystemy leśne
Prowadzone badania dotyczą określenia zmian poziomu wód podziemnych w środowiskach leśnych, takich jak: sosnowy bór bagienny, świerczyna bagienna, sosnowo-brzozowy las bagienny i ols oraz ich wpływu na stan drzewostanu (przyrosty i zdrowotność drzew) i występowanie rzadkich gatunków roślin, grzybów i zwierząt (Krzysztofiak, red. 2012b). Do szczegółowych celów prowadzonych badań należy: - zbadanie różnorodności gatunkowej grzybów, w tym porostów, oraz mszaków i owadów związanych z rosnącymi drzewami w wybranych zbiorowiskach leśnych; - ocena stanu zdrowotnego wybranych drzew w wytypowanych zbiorowiskach leśnych; - określenie zakresu zmian poziomu wód gruntowych na powierzchniach badawczych w różnych zbiorowiskach leśnych; - analiza powiązań zmian poziomu wód gruntowych z występowaniem wybranych grup organizmów oraz stanem zdrowotnym drzewostanu. Badania wybranych grup organizmów oraz zdrowotności drzew prowadzone były na 16 powierzchniach badawczych (po 4 w sosnowym borze bagiennym, sosnowo-brzozowym lesie bagiennym, świerczynie bagiennej i olsie), na których zainstalowane zostały piezometry. W celu prowadzenia badań zmian poziomu wód gruntowych na każdym stanowisku badawczym zainstalowano piezometr, w którym umieszczono ceramiczny czujnik ciśnienia i rejestrator zmian poziomu wody i temperatury wody. Łącznie zainstalowano 16 piezometrów zlokalizowanych w czterech zbiorowiskach leśnych, po 4 w każdym.
5.2.1. Powierzchnie badawcze
Badania prowadzone są na 16 stanowiskach zlokalizowanych w czterech typach lasów: w sosnowo-brzozowym lesie bagiennym, w sosnowym borze bagiennym, w świerczynie bagiennej oraz w olsie. Na każdym stanowisku zainstalowano piezometr oraz wytypowano i oznaczono po 5 drzew do badań. Stanowiska w sosnowo-brzozowym lesie bagiennym Thelypteridi-Betuletum pubescentis zlokalizowane zostały zarówno w części północnej Wigierskiego Parku Narodowego, w pobliżu doliny Wiatrołuży (oddz. 20), w jego części środkowej (odz. 129 i dz. ewid. 174), jak i w części południowej, w okolicy jeziora Krusznik (oddz. 217). Stanowiska w sosnowym borze bagiennym Vaccinio uliginosi-Pinetum zlokalizowane zostały zarówno w części północnej Wigierskiego Parku Narodowego (oddz. 22), w jego części środkowej zachodniej (odz. 111) oraz wschodniej (oddz. 233), jak i w części południowej na Suchym Bagnie (oddz. 276). Stanowiska w świerczynie bagiennej Sphagno girgensohnii-Piceetum zlokalizowane zostały w środkowej części Wigierskiego Parku Narodowego, od strony zachodniej (oddz. 111, 114 i 121) oraz od strony wschodniej (oddz. 206). Stanowiska w olsie porzeczkowym Ribo nigri-Alnetum zlokalizowane zostały w części północnej Wigierskiego Parku Narodowego, w pobliżu doliny Wiatrołuży (oddz. 25) oraz w części środkowej zachodniej (odz. 86, 117 i 122).
5.2.2. Wyniki badań
5.2.2.1. Inwentaryzacja organizmów na powierzchniach badawczych
Owady W trakcie badań na pniach drzew kontrolnych stwierdzono obecność jedynie dwóch gatunków owadów: Agelastica alni i Microbregma emarginatum. Pierwszy z nich Agelastica alni, to pospolity palearktyczny gatunek występujący na terenie całej Polski (Burakowski i in. 1991) (Fot. 15). Jest to foliofag wykazujący tendencje do masowych pojawów, odżywiający się liśćmi olszy. Wyrządza szkody w młodnikach osłabiając drzewa, jednak w przypadku starszych drzewostanów olszowych ma niewielkie znaczenie i nie wpływa znacząco na ich kondycję. U samicy tego gatunku cykl składania jaj zaczyna się po przezimowaniu i trwa aż do połowy lipca, potrafi ona złożyć w tym czasie około 600-900 jaj. Larwy spotyka się na liściach do początku września. Przepoczwarczenie odbywa się w glebie w okresie od lipca do września (Szujecki 1995).
Fot. 15. Agelastica alni na pniu olszy
Drugi gatunek Microbregma emarginatum to środkowoeuropejski gatunek występujący najprawdopodobniej na terenie całej Polski. Owad ten roi się w maju, a postacie doskonałe można spotkać do końca lipca. Larwy tego chrząszcza żerują w zewnętrznej warstwie korowiny starych świerków. (Burakowski i in. 1986). Otwory wylotowe tego kołatka na powierzchni kory są często mylone z otworami wylotowymi niektórych korników, będących groźnymi szkodnikami świerka. Żer tego gatunku nie wpływa na kondycje i zdrowotność drzew, ani nie obniża wartości surowca drzewnego. Dlatego można przyjąć, że nie ma żadnego negatywnego wpływu na drzewostan. (Dominik, Starzyk 1989). Na pomocniczych powierzchniach badawczych stwierdzono obecność jedenastu taksonów owadów: Callidium violaceum (Fabricius, 1775),Elateroides dermestoides (Linnaeus, 1761), Ips typographus (Linnaeus, 1758), Rhagium inquisitor (Linnaeus, 1758), Saperda carcharias (Linnaeus, 1758), Scolytus ratzeburgii Janson, 1856, Tetropium castaneum (Linnaeus, 1758), Tomicus piniperda (Linnaeus, 1758), Monochamus sp., Tremex fuscicornis (Fabricius, 1787) i Xiphydria camelus (Linnaeus, 1758).
Grzyby W trakcie przeprowadzonych w 2013 roku badań terenowych, na 80 okazach drzew, dokonano łącznie 126 notowań grzybów. W tym czasie stwierdzono występowanie ok. 40 taksonów grzybów makroskopijnych związanych bezpośrednio lub pośrednio z analizowanymi składnikami dendroflory. Wśród zebranych grzybów zidentyfikowano do chwili obecnej 29 gatunków, w tym 28 przedstawicieli grzybów podstawkowych i 1 przedstawiciela grzybów workowych.
Fot. 16. Podophacidium xanthomelum (A), Mycena pseudocorticola (B)
Wśród przedstawicieli macromycetes brak jest gatunków chronionych, natomiast znajdują się wśród nich grzyby zamieszczone na „Czerwonej liście grzybów wielkoowocnikowych w Polsce” (Wojewoda, Ławrynowicz 2006). Są to dwa gatunki narażone (kategoria zagrożenia – V), tj.:Mycena pseudocorticola (Fot. 16B) i Mycena purpureofusca oraz 2 gatunki grzybów rzadkich (kategoria zagrożenia – R), tj.: Entoloma rhodocalix i Galerina triscopa. Na badanych powierzchniach w 2013 r. znaleziono 33 taksony porostów. W stosunku do roku 2012 znaleziono 4 nowe taksony: Ochlolechia sp., Opegrapha sp., Placynthiella dasaea i Trapeliopsis granulosa. Są to wszystko porosty skorupiaste stwierdzone na pojedynczych stanowiskach, a znalezione plechy są małe i młode – najprawdopodobniej porosty te zasiedliły badane drzewa w ciągu ostatniego roku. Nie odnaleziono jednego gatunku – Ropalospora viridis – w 2012 r. występował on w postaci małych skorupiastych plech na pniach dwóch sosen; nie można wykluczyć, że plechy odpadły z korą, która łuszczy się na tych drzewach w sposób naturalny.
Mszaki W wyniku przeprowadzonych badań, na 80 przeanalizowanych drzewach stwierdzono występowanie 54 taksonów mszaków, tj. 14 gatunków wątrobowców oraz 38 mchów. Ponadto stwierdzono obecność darni z rodzaju Ulota i Orthotrichum, ale ich identyfikacja do gatunku, była niemożliwa, głównie ze względu na brak sporogonów oraz niewielkie rozmiary okazów. W porównaniu z zeszłorocznymi badaniami, flora epifityczna badanych drzew zubożała o dwa gatunki: Jamesoniella autumnalis oraz Leucobryum glaucum. Oba taksony występowały na szyi korzeniowej badanych obiektów. Jednocześnie musko- i hepatikoflora badanych drzew wzbogaciła się aż o pięć taksonów: Calypogeia azurea, Calypogeia integristipula, Odontoschisma denudatum, Orthotrichum sp. oraz Plagiomnium elipticum. Brioflora porastająca pnie badanych drzew jest dość uboga. W sumie, we wszystkich fitocenozach odnotowano 20 gatunków, w tym trzy taksony to epifity obligatoryjne. Są to: Radula complanata, Ulota sp. oraz Orthotrichum sp. Dwa razy bogatsza jest flora mchów i wątrobowców porastająca szyje korzeniowe. Mszaki występujące na tym typie mikrosiedliska określane są jako epifityczno-naziemne. Duży udział mają tu gatunki przechodzące z dna lasu, a więc im bogatsza flora dna lasu, tym większe bogactwo gatunkowe u nasady drzew. Zaskakujące było pojawienie się na szyjach korzeniowych np. Geocalyx graveolens, który jest gatunkiem występującym na wilgotnym humusie, czy Calypogeia sphagnicola, gatunku typowo torfowiskowego. Procent pokrycia kory pni drzew przez epifity był zazwyczaj niewielki. Większe powierzchnie porastały tylko Hypnum cupressiforme, Orthodicranum montanum oraz Platygyrium repens, pozostałe taksony tworzyły raczej małe darnie. Na szyjach korzeniowych z wysokim pokryciem notowano m.in. Eurhynchium angustirete, Hylocomium splendens, Hypnum cupressiforme (Fot. 17), Lepidozia reptans, Orthodicranum flagellare, Orthodicranum montanum i Platygyrium repens.
Fot. 17. Szyja korzeniowa pokryta gęstym kobiercem Hypnum cupressiforme
W porównaniu z 2012 r. nie udało się potwierdzić 30 notowań (wystąpień) mszaków, jednocześnie w 2013 r. zarejestrowano aż 50 nowych wystąpień. Ostateczny bilans wyraźnie wskazuje na tendencję do zwiększania (choć nieznacznego) liczby darni epifitów. W odniesieniu do zdecydowanej liczby gatunków nie udało sie potwierdzić pojedynczych notowań. Wyjątek stanowią dwa gatunki: Dicranum scoparium i Pohlia nutans, w przypadku których nie udało się potwierdzić po cztery notowania. Wzrost liczby notowań dotyczy zarówno mchów jak i wątrobowców. Najwięcej nowych darni zaobserwowano w przypadku Lophocolea heterophylla, której bilans notowań wzrósł o pięć. Brioflora epifityczna analizowanych drzew nie jest zbyt zróżnicowana. Znaczna część gatunków jest taka sama we wszystkich zbiorowiskach, jednak poszczególne zbiorowiska różnią się nie tylko liczbą gatunków, ale także częstością ich występowania. Najuboższe w epifity są drzewa w sosnowym borze bagiennym. W sumie stwierdzono tu tylko dziewięć gatunków. Najczęściej notowanym epifitem był Pleurozium schreberi. Zdecydowaną większość drzew w tym zbiorowisku stanowią sosny, których łuszcząca kora zdecydowanie nie sprzyja rozwojowi brioflory epifitycznej. Jedynie u nasady szyi korzeniowych znajdywano darnie wątrobowców takich jak: Calypogeia muelleriana, Cephalozia connivens, Lophocolea bidentata i Odontoschisma denudatum. Znacznie bogatsza jest brioflora epifityczna w sosnowo-brzozowym lesie bagiennym, która łącznie liczy 20 taksonów. Najczęściej notowanymi epifitami są tu Lepidozia reptans, Lophocolea heterophylla, Orthodicranum montanum, Plagiothecium laetum, Ptilidium pulcherimmum i Tetraphis pellucida. Nieco bogatsze gatunkowo w epifity są drzewa w świerczynie bagiennej. W zbiorowisku tym w sumie odnaleziono 31 gatunków, spośród których najwięcej notowań miały: Hypnum cupressiforme, Lophocolea heterophylla, Orthodicranum montanum, Plagiothecium laetum i ortotropowy Tetraphis pellucida. Najbogatszą florą epifityczną cechują się drzewa w olsie. Najczęściej notowane mszaki to Herzogiella seligeri, Hypnum cupressiforme, Lophocolea heterophylla, Orthodicranum montanum, Plagiothecium laetum oraz Platygyrium repens. W porównaniu z danymi z 2012 r. największe zmiany zaszły w brioflorze epifitycznej w świerczynie bagiennej. W zbiorowisku tym liczba notowań mszaków zmniejszyła się o dziewięć, a jednocześnie kilkanaście gatunków zwiększyło swój zasięg, o czym świadczy aż 21 nowych notowań. W pozostałych zbiorowiskach różnice w liczbie wystąpień gatunków są bardzo małe. W borze bagiennym dwa notowania ubyły, pięć przybyło, w sosnowo-brzozowym lesie bagiennym osiem ubyło, 12 przybyło, jedynie w olsie 12 ubyło, a tylko osiem wystąpień przybyło. Flora mchów i wątrobowców badanych drzew należy do stosunkowo ubogich. Większość gatunków to taksony częste, o szerokiej amplitudzie ekologicznej. Zdecydowanie zaznacza się minimalny udział obligatoryjnych epifitów. Na wszystkich przebadanych drzewach stwierdzono ich tylko trzy. Wśród zanotowanych gatunków epifitycznych 13 to gatunki objęte ochroną, przy czym w większości są to pospolite gatunki borowe, które przeszły z dna lasu na szyje korzeniowe drzew. Pozytywnym zjawiskiem jest pojawienie się kolejnej darni Ulota sp. Jeszcze do niedawna liczba stanowisk gatunków z tego rodzaju, podobnie jak wielu obligatoryjnych epifitów, w szybkim tempie zmniejszała się w całej Europie. Za główny czynnik powodujący ich wymieranie uznano zanieczyszczenie powietrza, głównie wzrost stężenia związków siarki i azotu (Barkmann 1958; Dierssen 2001). Tymczasem od lat 90-tych, obserwuje się wzrost liczby stanowisk mchów epifitycznych, zwłaszcza taksonów z rodziny Orthotrichaceae (Plàsek 2007; Stebel 2006, 2010; Fudali 2012).
5.3. Monitoring inwazyjnych gatunków roślin obcego pochodzenia
Celem podjętych badań jest ocena zagrożenia rodzimej flory (różnorodności gatunkowej) i siedlisk przyrodniczych zlewni badawczej ze strony inwazyjnych gatunków roślin obcego pochodzenia oraz ocena zmian (ich tendencji i kierunków) populacji tych gatunków. Badania prowadzone były na terenie całej zlewni badawczej oraz jej otuliny przez cały sezon wegetacyjny, przy wykorzystaniu odbiorników GPS, a uzyskane wyniki naniesione zostały na mapę numeryczną obszaru. Wyniki rozmieszczenia i wielkości populacji poszczególnych gatunków analizowano na tle warunków siedliskowych. Z założenia badania skupiały się nie tylko na określeniu lokalizacji występowania i tempa rozprzestrzeniania się obcych gatunków, ale obejmowały również zagadnienia związane z biologią i ekologią tych gatunków oraz ich wymaganiami siedliskowymi.
5.3.1. Wyniki badań
W 2013 roku szczególną uwagę zwrócono na występowanie niecierpka gruczołowatego Impatiens glandulifera - jednego z najbardziej inwazyjnych gatunków w ostatnich latach. Wstępne badania przeprowadzone na terenie Wigierskiego Parku Narodowego wykazały, że gatunek ten zagraża przede wszystkim najbardziej naturalnym środowiskom przyrodniczym, jakimi są doliny rzeczne. Niecierpek gruczołowaty jest rośliną jednoroczną, występującą w stanie dzikim w Azji Środkowej, w rejonie Himalajów oraz w zachodnich Indiach i Pakistanie. Zawleczony do Polski jest dość często uprawiany jako roślina ozdobna. Zadomowił się w dolinach rzek i strumieni, na obrzeżach wilgotnych lasów, zwłaszcza olsów i łęgów - najchętniej zasiedla podmokłe, okresowo zalewane tereny bogate w związki organiczne. Niecierpek gruczołowaty rozprzestrzenia się tylko poprzez nasiona. Jedna roślina może wyprodukować do 4000 nasion, umieszczonych w podłużnych torebkach, pękających przy dotknięciu i wystrzeliwujących nasiona nawet na odległość 7 metrów. Dzięki dużej produkcji nasion, ich synchronicznemu wzejściu oraz szybkiemu wzrostowi roślina ta skutecznie konkuruje z naszymi rodzimymi gatunkami i z łatwością wypiera naturalną roślinność, tworząc jednogatunkowe łany o znacznej powierzchni (Fot. 18).
Fot. 18. Niecierpek gruczołowaty Impatiens glandulifera
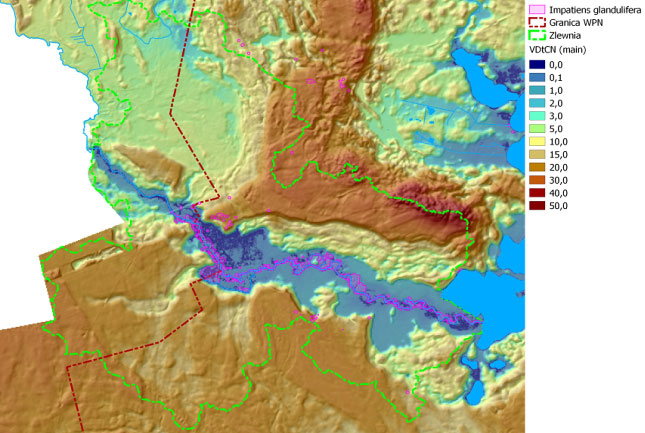
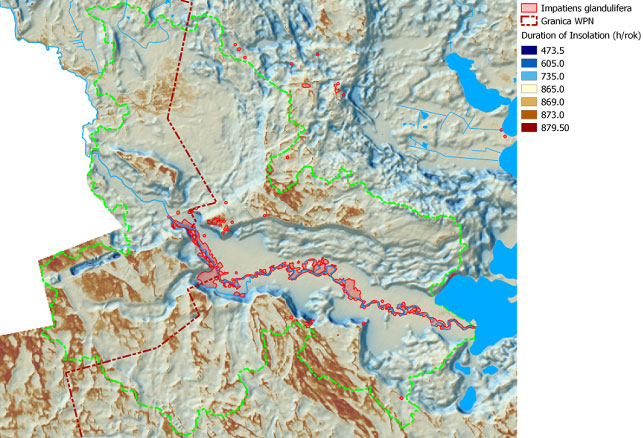
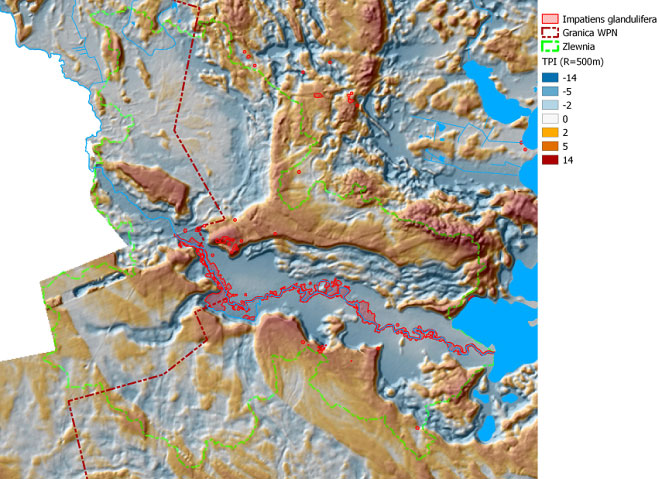
W wyniku przeprowadzonych w 2013 roku badań, w dolinie Czarnej Hańczy określona została szczegółowa lokalizacja niecierpka gruczołowatego. Lokalizację stanowisk tego gatunku na obszarze zlewni badawczej przedstawiają rysunki: rys. 101 - na tle obszarów podmokłych, rys. 102 - na tle obszarów o zróżnicowanym czasie nasłonecznienia, rys. 103 - na tle obszarów wyniesionych i obniżonych. Wyniki uzyskiwane z monitoringu inwazyjnych gatunków roślin obcego pochodzenia wykorzystywane są do planowania zabiegów czynnej ochrony rodzimej przyrody, w tym różnorodności biologicznej. Na obszarze zlewni badawczej WIGRY na tej podstawie wykonywane są prace związane z ograniczeniem wielkości populacji niecierpka gruczołowatego. Przy pomocy środków Instrumentu Finansowego LIFE+ Unii Europejskiej oraz Narodowego Funduszu Ochrony Środowiska i Gospodarki Wodnej, na powierzchni kilkunastu hektarów rozpoczęto zabiegi usuwania roślin tego gatunku, które będą prowadzone przez 4 kolejne lata.
Rys. 101. Mapa obrazująca rozmieszczenie niecierpka gruczołowatego na tle obszarów podmokłych – obszary te zostały wyznaczone na podstawie współczynnika VDtCN (Vertical Distance to Chanell Network)
Rys. 102. Mapa obrazuje rozmieszczenie niecierpka gruczołowatego na tle obszarów o zróżnicowanym czasie nasłonecznienia (h/rok)
Rys. 103. Mapa obrazuje rozmieszczenie niecierpka gruczołowatego na tle obszarów wyniesionych i obniżonych - układ tych obszarów został wyznaczony na podstawie współczynnika TPI (Terrain Position Index) dla kołowych powierzchni o promieniu 200 m
Niezwykle ważnym elementem badań jest monitoring efektów ekologicznych podejmowanych zabiegów zwalczania niecierpka gruczołowatego. Jego wyniki pozwalają ocenić skuteczność działań oraz opracować najlepszą metodę zwalczania gatunku. Już pierwsze wyniki prowadzonych badań wskazują, że kluczowymi zagadnieniami w ograniczaniu populacji niecierpka gruczołowatego są: sposób usuwania roślin (wyrywanie, koszenie), powtarzalność zabiegów usuwania roślin w tym samym sezonie oraz terminy prowadzenia tych zabiegów. Podstawowym założeniem prowadzenia zabiegów musi być wyeliminowanie z gleby jak największej liczby nasion niecierpka, a nie tylko usunięcie rośliny. Niecierpek gruczołowaty skutecznie konkuruje z rodzimymi gatunkami roślin tworząc zwarte łany uniemożliwiające im wzrost. Przeprowadzając zabiegi usuwania niecierpka w odpowiednich terminach i z odpowiednią częstotliwością można znacząco ograniczyć wielkość jego populacji i dać szanse na odtworzenie się rodzimej flory.
Fot. 19. Teren opanowany niemal wyłącznie przez niecierpka gruczołowatego
Fot. 20. Niecierpek gruczołowaty może dorastać do 3 m wysokości
Fot. 21. Po kilkukrotnym przeprowadzeniu zabiegu zwalczania niecierpek gruczołowaty występuje na powierzchni sporadycznie
Po jednym sezonie prowadzenia zabiegów usuwania niecierpka gruczołowatego, z odpowiednią częstotliwością i w odpowiednich terminach, znacząco ograniczono występowanie tego inwazyjnego gatunku rośliny. Pozwoliło to na pojawienie się na tym terenie roślin rodzimych, które wcześniej tu występowały.
Literatura Barkmann J. J. 1958. Phytosociology and ecology of cryptogamic epiphytes. Van Gorcum & Comp. N. V. G. H. Hak & Dr H. J. Prakke, Assen. Burakowski B., Mroczkowski M., Stefańska J. 1986: Kat. Fauny Pol. Część XXIII, Tom 11. Chrząszcze Coleoptera, Dermestoidea, Bostrichoidea, Cleroidea i Lymexyloidea. PWN, Warszawa. 243 ss. Burakowski B., Mroczkowski M., Stefańska J., 1991: Kat. Fauny Pol. Część XXIII, Tom 17. Chrząszcze Coleoptera, Stonkowate – Chrysomelidae, cz 2. PWN, Warszawa. 227 ss. Cieśliński S., Czyżewska K., Fabiszewski J. 2003. Czerwona lista porostów wymarłych i zagrożonych w Polsce. Monographiae Botanicae 91: 13–49. Dierssen K. 2001. Distribution, ecological amplitude and phytosociological characterization of European bryophytes. Bryophyt. Biblioth. 56: 1-289. Dominik J., Starzyk j. R., 1989: Owady niszczące drewno. PWRiL. Warszawa. 524 ss. Fałtynowicz W. 2003. The lichens, lichenicolous and allied fungi of Poland – an annotated checklist. Instytut Botaniki im. W. Szafera, Polska Akademia Nauk, Kraków, 435 ss. Fałtynowicz W. 2013. Raport z badań mszaków w ramach realizacji projektu „Badania dynamiki zmian poziomu wód podziemnych oraz ich wpływu na podmokłe ekosystemy leśne Wigierskiego Parku Narodowego - etap II. Porosty”. Maszynopis, WPN. Fałtynowicz W. 2013. Raport z realizacji badań nad mszakami w ramach projektu „Badania organizmów saproksylicznych w różnych ekosystemach leśnych Wigierskiego Parku Narodowego – etap II. Porosty”. Maszynopis, WPN. Fudali E. 2012. Recent tendencies in distribution of epiphytic bryophytes in urban areas: a Wrocław case study (south-west Poland). Pol. Bot. Journ. 57(1): 231-241. Halama M. 2013. Raport z badań mszaków w ramach realizacji projektu „Badania dynamiki zmian poziomu wód podziemnych oraz ich wpływu na podmokłe ekosystemy leśne Wigierskiego Parku Narodowego - etap II. Grzyby wielkoowocnikowe”. Maszynopis, WPN. Halama M. 2013. Raport z realizacji badań nad mszakami w ramach projektu „Badania organizmów saproksylicznych w różnych ekosystemach leśnych Wigierskiego Parku Narodowego – etap II. Grzyby wielkoowocnikowe”. Maszynopis, WPN. Kepel A., Fałtynowicz W., Zalewska A., Kujawa A. 2012. Aktualizacja listy gatunków grzybów objętych ochroną gatunkową oraz wskazania dla ich ochrony. Mscr., Poznań. Klama H. 2002. Distribution patterns of liverworts (Marchantiopsida) in natural forest communities (Białowieża Primeval Forest, NE Poland). – University of Bielsko-Biała, Bielsko-Biała, ss. xiv + 278. Krzysztofiak L. (red.), Fałtynowicz W., Panek E., Staniaszek-Kik M., Halama M., Sawoniewicz M., Romański M., Krzysztofiak A., Janecki T., Mackiewicz A. 2012. Badania organizmów saproksylicznych w różnych ekosystemach leśnych Wigierskiego Parku Narodowego (maszynopis). WPN Krzywe. Kukwa M. 2005. The lichen species Cladonia incrassata (Lecanorales, Ascomycota lichenisati) in Poland, and notes on C. anitae. – Polish Bot. J. 50.1: 69–73. Muhle H., LeBlanc F. 1975. Bryophyte and lichen succession on decaying logs. I. Analyses along on evaporational gradient in eastern Canada. – J. Hattori Bot. Lab. 39: 1-33. Panek E., Romański M. 2013. Raport z realizacji badań nad mszakami w ramach projektu „Badania organizmów saproksylicznych w różnych ekosystemach leśnych Wigierskiego Parku Narodowego – etap II. Śluzowce”. Maszynopis, WPN. Plášek V. 2007. Gatunki z rodziny Orthotrichaceae (Bryophyta) i problemy ich ochrony. W: Kasza H., Klama H. (red.), Zapobieganie zanieczyszczeniu, przekształcaniu i degradacji środowiska 14: 137-140. Sawoniewicz M. 2013. Raport z badań mszaków w ramach realizacji projektu „Badania dynamiki zmian poziomu wód podziemnych oraz ich wpływu na podmokłe ekosystemy leśne Wigierskiego Parku Narodowego - etap II. Chrząszcze (Coleoptera)”. Maszynopis, WPN. Sawoniewicz M. 2013. Raport z realizacji badań nad mszakami w ramach projektu „Badania organizmów saproksylicznych w różnych ekosystemach leśnych Wigierskiego Parku Narodowego – etap II. Chrząszcze (Coleoptera)”. Maszynopis, WPN. Staniaszek-Kik M. 2013a. Raport z badań mszaków w ramach realizacji projektu „Badania dynamiki zmian poziomu wód podziemnych oraz ich wpływu na podmokłe ekosystemy leśne Wigierskiego Parku Narodowego - etap II. Mszaki”. Maszynopis, WPN. Staniaszek-Kik M. 2013s.Raport z realizacji badań nad mszakami w ramach projektu „Badania organizmów saproksylicznych w różnych ekosystemach leśnych Wigierskiego Parku Narodowego – etap II. Mszaki”. Maszynopis, WPN. Stebel A. 2010. Wpływ zbiorników zaporowych na Dunajcu w Pieninach na florę mchów tego regionu. Pieniny – Zapora – Zmiany – Monografie Pienińskie 2: 161–171. Stebel A. 2006. The mosses of the Beskidy Zachodnie as a paradigm of biological and environmental changes in the flora of the Polish Western Carpathians. Habilitation Thesis 17/2006. Medical University of Silesia in Katowice, Sorus, Katowice–Poznań. Szujecki A., 1995: Entomologia leśna. Tom II.Wydawnictwo SGGW. Warszawa. 408 ss. Wojewoda W., Ławrynowicz M. 2006. Red list of the macrofungi in Poland. (w:) Mirek Z., Zarzycki K., Wojewoda W., Szeląg Z., (red.). Red list of plants and fungi in Poland. W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, p. 55–70.
|
||||||