kod siedliska: 7110, siedlisko priorytetowe
*Torfowiska wysokie z roślinnością torfotwórczą (żywe)
Definicja
Otwarte mszary na skrajnie ubogich w związki odżywcze, bardzo kwaśnych i silnie wilgotnych torfach, zasilane wyłącznie lub niemal wyłącznie przez wody opadowe i przez to wybitnie uzależnione od cech klimatu. Lustro wody w złożu torfowym jest położone wyżej w stosunku do poziomu wody gruntowej w otoczeniu torfowiska. Zbiorowiska roślinne torfowisk wysokich budowane są przez bardzo nieliczną, ekologicznie bardzo wyspecjalizowaną grupę roślin, głównie torfowce, krzewinki, zielne byliny o trawiastym pokroju, sporadycznie gatunki krzewiaste i drzewiaste. Torfowiska wysokie cechuje makro- i mikromorfologiczne zróżnicowanie powierzchni złoża torfu i odpowiadające temu jakościowe i przestrzenne zróżnicowanie siedlisk i roślinności. Fitocenozy należą do różnych klas zbiorowisk. Najbardziej torfotwórczy charakter mają specyficzne dla torfowisk wysokich zbiorowiska z klasy Oxycocco-Sphagnetea, porastające mikrosiedliska określane jako kępki. Narastanie złoża torfowego kończy się, gdy bilans wodny torfowiska (dopływ i odpływ) zostanie zrównoważony.
Charakterystyka
Torfowiska wysokie w sensie ekologicznym należą do siedlisk skrajnych: cechuje je stałe wysokie uwilgocenie, silnie kwaśny odczyn (pH 3,5 - 4,5), wyjątkowo niska trofia. Warunki takie powstają w wyniku całkowitego odizolowania przez warstwę torfu powierzchni torfowiska od wpływu wód gruntowych lub powierzchniowych i pełne uzależnienie roślinności od wody pochodzącej z opadów atmosferycznych. W pierwszym etapie jest ona retencjonowana w specjalnych komórkach torfowców, następnie trwale zmagazynowana w pokładzie torfu i może stanowić do 97% jego świeżej masy. Dzięki temu torfowiska wysokie w stosunku do otoczenia stanowią odrębny, niezależny układ hydrologiczny. W sensie funkcjonalnym żywe torfowisko składa się z dwu warstw: akrotelmu i katotelmu. Akrotelm, czyli warstwa powierzchniowa, położona powyżej przeciętnego stanu wody w torfowisku, jest warstwą torfotwórcza. Zachodzi w niej proces mikrobiologicznego rozkładu obumarłej masy roślinnej z aktualnie występującej roślinności. Katotelm stanowi warstwa martwego torfu, stale wysyconego wodą i przez to niepodlegającego dalszemu rozkładowi. Obniżenie poziomu wody w torfowisku zakłóca równowagę między obiema warstwami, w skrajnym przypadku prowadzi do całkowitego zaniku akrotelmu i zmniejszania się objętości katotelmu poprzez rozkład zmagazynowanego w nim torfu.
Powierzchnia torfowiska wysokiego jest mniej lub bardziej wypukła, przez co w granicach całego torfowiska zaznacza się zróżnicowanie wilgotnościowe i troficzne. W zwykle płaskiej, najwyżej położonej i najbardziej wilgotnej części kopuły -wierzchowinie - dodatkowo występuje wyraźna mikrorzeźba, w postaci zespołu kęp i dolinek, ze swoistymi zbiorowiskami roślinnymi. Na nachylonych, suchszych zboczach mogą rosnąć drzewa, a na granicy torfowiska i mineralnego otoczenia występuje silnie podtopiony, minerotroficzny okrajek, do którego spływa woda z kopuły oraz terenu otaczającego torfowisko. Na rosnących torfowiskach lustro wody występuje w pobliżu ich powierzchni i wykazuje niewielkie wahania w ciągu roku. Układa się ono zgodnie z kształtem sylwetki całego złoża torfowego, a w przypadku jego rozcięcia - dostosowane jest do poszczególnych fragmentów. Powoduje to, że rozmieszczenie roślinności torfotwórczej i akumulacja torfu ograniczone są do najlepiej uwilgoconych partii złoża.
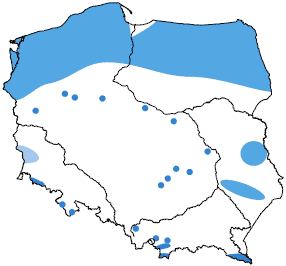
Optymalne warunki do wykształcania się torfowisk wysokich występują w strefie klimatu umiarkowanego i wilgotnego. W Polsce ten typ torfowisk występuje przede wszystkim na północy, w pasie przymorskim, gdzie duże torfowiska kopułowe osiągają regionalną południową granicę zasięgu. Ponadto nieliczne torfowiska wysokie rozmieszczone są w środkowej części kraju, w górach i na Podhalu. Torfowiska wysokie rozwijają się na obszarach wododziałowych, ale również w obniżeniach dolinnych i pradolinach, zawsze jednak poza strefą oddziaływania wód gruntowych czy zalewowych. Największe z nich, tzw. torfowiska kopułowe typu bałtyckiego, mają powierzchnię dochodzącą nawet do 200 ha, lecz jest ich tylko około 60. Znacznie więcej jest niewielkich (1-10 ha) złóż w bezodpływowych zagłębieniach terenu. Łącznie torfowiska wysokie w Polsce stanowią zaledwie 4,3% ogółu powierzchni krajowych torfowisk. Szacuje się, że żywe torfowiska wysokie występują obecnie na zaledwie 1 % swojego pierwotnego areału.
Podział na podtypy
Torfowiska wysokie, jako jeden z trzech szeroko ujmowanych typów torfowisk, w granicach całego swojego europejskiego obszaru występowania wykazują wyraźne zróżnicowanie sylwetki całego złoża i występującej na nim roślinności. Jest ono skorelowane ze zróżnicowaniem klimatu -wygasaniem ku wschodowi cech oceanicznych, a narastaniem cech kontynentalnych, a także z gradientem termicznym związanym z szerokością geograficzną i wyniesieniem terenu nad poziom morza. Geograficzne położenie i rzeźba terenu Polski powodują, że między poszczególnymi regionami w kraju istnieją wyraźne różnice klimatyczne, w efekcie czego na torfowiskach wysokich, oprócz gatunków wspólnych, rosną także gatunki typowe dla atlantyckiej, borealnej lub kontynentalnej części Europy. Powoduje to florystyczno-geograficzne zróżnicowanie krajowych zbiorowisk mszarnych, które jest podstawą poniższego, regionalnego podziału na podtypy:
*7110-1 Niżowe torfowiska wysokie
*7110-2 Sudeckie torfowiska wysokie
*7110-3 Karpackie torfowiska wysokie
Umiejscowienie siedliska w polskiej klasyfikacji fitosocjologicznej
Funkcjonujące w sensie ekologicznym (żywe) torfowisko wysokie, jako typ siedliska, jest złożonym układem ekologiczno-roślinno-przestrzennym. Zróżnicowanie warunków siedliskowych w jego granicach jest na tyle znaczące, że gatunki tworzą nie tylko różne zespoły, ale zespoły te należą do różnych klas zbiorowisk jak: Oxycocco-Sphagnetea, Scheuchzerio-Caricetea nigrae, Utricularietea intermedio-minoris, w bardzo ograniczonym zakresie również Vaccinio-Piceetea. Spośród nich tylko zespoły klasy Oxycocco-Sphagnetea są specyficzne dla torfowisk wysokich i w najpełniejszym zakresie odpowiadają ekologicznej definicji siedliska jako typu, natomiast w skali wewnętrznego zróżnicowania siedliska reprezentują roślinność mikrosiedlisk, jakim są kępki. Zespoły roślinne pozostałych klas i niższych jednostek w ich obrębie odnoszą się do takich elementów kompleksu przestrzennego, jak: dolinki (Scheuchzerietalia palustris), zbocze kopuły (Vaccinio-Piceetea), okrajek (Utricularietea intermedio-minoris, Scheuchzerietalia palustris, Caricetalia nigrae).
Klasyfikacja fitosocjologiczna specyficznych dla torfowisk wysokich mszarów kępowych w skali Europy jest ciągle przedmiotem dyskusji, co wynika m.in ze stosowania różnych kryteriów podziału i uzupełniania wiedzy o dane z regionów dotąd słabo lub w ogóle niebadanych. Dotyczy to również Polski. Inwentarz krajowych zbiorowisk występujących na torfowiskach wysokich, zawarty w syntetycznych opracowaniach, nie uwzględnia wszystkich zespołów zidentyfikowanych wtoku dotychczasowych badań. Brak jest również całościowej rewizji materiału oraz jego analizy w aspekcie geograficznym, odniesionym do całego europejskiego obszaru występowania i zróżnicowania mszarnych ombrotroficznych zbiorowisk roślinnych. Poniższy wykaz ma charakter tymczasowy i obejmuje zespoły i zbiorowiska roślinne zidentyfikowane na krajowych torfowiskach wysokich bezdrzewnych oraz z niewielkim udziałem drzew, które jednak nie nadają fitocenozom struktury lasu. W nawiasach podano najczęściej spotykane synonimy oraz inne podobne opisywane zespoły. Występowanie zespołów oznaczonych „? " wymaga weryfikacji, jednak jest prawdopodobne na terenie Polski (stwierdzono je na torfowiskach górskich naszych południowych sąsiadów).
Zespoły kępkowe (kod Physis 51.11):
Klasa Oxycocco-Sphagnetea zbiorowiska mokrych wrzosowisk i torfowisk wysokich
Rząd Erico-Sphagnetalia (Sphagno-Ericetalia)
atlantyckie zbiorowiska mokrych wrzosowisk i torfowisk wysokich
Związek Oxycocco-Ericion zbiorowiska torfowisk wysokich
oceanicznej i suboceanicznej części Europy
Zespoły i zbiorowiska:
Erico-Sphagnetum magellanici mszar wysokotorfowiskowy z wrzoścem bagiennym
Scirpo austriaci-Sphagnetum papillosi (fragmentarycznie)
zespół wełnianeczki darniowej i torfowca brodawkowanego
Sphagnum papillosum zbiorowisko torfowca brodawkowatego
Rząd Sphagnetalia magellanici mszarne zbiorowiska kępowe torfowisk wysokich
środkowej i borealnej części Europy
Związek Sphagnion magellanici środkowoeuropejskie mszary wysokotorfowiskowe
Zespoły:
Sphagnetum magellanici mszar kępowy z torfowcem magellańskim
Eriophoro vaginati-Sphagnetum recurvi (=zbiorowisko Eriophorum vaginatum-Sphagnum fallax)
zespół wełnianki pochwowatej i torfowca kończystego
Eriophoro-Trichophoretum caespitosi zespół wełnianki pochwowatej i wełnianeczki darniowej
Ledo-Sphagnetum magellanici zespół bagna zwyczajnego i torfowca magellańskiego
? Trichophorum alpinum-Sphagnum compactum (= Sphagneto-Trichophoretum alpini)
zespół wełnianeczki alpejskiej i torfowca szorstkiego
Związek Oxycocco (microcarpi)-Empetrion hermaphroditi
subarktyczno-borealne mszary wysokotorfowiskowe
Zespoły:
Empetro-Trichophoretum austriaci zespół bażyny obupłciowej i wełnianeczki darniowej
Sphagno robusti-Empetretum hermaphroditi zespół torfowca Russowa i bażyny obupłciowej
Chamaemoro-Empetretum hermaphroditi zespół maliny moroszki i bażyny obupłciowej
Empetro hermaphroditi-Sphagnetum fusci zespół bażyny obupłciowej i torfowca brunatnego
Zespoły dolinkowe (Kod Physis 51.12):
Klasa Scheuchzerio-Caricetea nigrae zbiorowiska torfowisk niskich, przejściowych i dolinek
na torfowiskach wysokich
Rząd Scheuchzerietalia palustris torfotwórcze zbiorowiska torfowisk przejściowych i dolinek
na torfowiskach wysokich
Związek Rhynchosporion albae mszary przejściowo-torfowiskowe i dolinkowe
Zespoły:
Caricetum limosae (oraz inne opisywane zespoły): Sphagno lindbergii-Caricetum limosae,
Sphagno duseni-Caricetum limosae, Scheuchzerio-Sphagnetum cuspidati) mszar dolinkowy
z turzycą bagienną
Rhynchosporetum albae mszar przygiełkowy
Eriophoro angustifolii-Sphagnetum recurvi (oraz inne opisywane zespoły:
Calliergo sarmentosi-Eriophoretum angustifolii) zespół wełnianki wąskolistnej
i torfowca kończystego
Zespoły okrajkowe (Kod Physis 51.15):
Klasa Scheuchzerio-Caricetea nigrae zbiorowiska torfowisk niskich, przejściowych
i dolinek na torfowiskach wysokich
Rząd Scheuchzerietalia palustris torfotwórcze zbiorowiska torfowisk przejściowych
i dolinek na torfowiskach wysokich
Związek Rhynchosporion albae mszary przejściowo-torfowiskowe i dolinkowe
Zespół
Eriophoro angustifolii-Sphagnetum recurvi (oraz inne opisywane zespoły: Calliergo sarmentosi-
Eriophoretum angustifolii) zespół wełnianki wąskolistnej i torfowca kończystego
Związek Caricion lasiocarpae zbiorowiska wąskolistnych turzyc na silnie kwaśnych torfowiskach
przejściowych, o subborealnym typie rozmieszczenia
Zespoły:
Caricetum lasiocarpae (w tym: podzespół ze Sphagnum fallax = Carici filiformis-
Sphagnetum apiculati) zespół turzycy nitkowatej
Caricetum rostratae (w tym: podzespół ze Sphagnum fallax = Carici rostratae-
Sphagnetum apiculati oraz inne: Carici rostratae-Drepanocladetum fluitantis)
zespół turzycy dziobkowatej
Rząd Caricetalia nigrae kwaśne młaki niskoturzycowe
Związek Caricion nigrae kwaśne młaki niskoturzycowe
Zespoły:
Carici echinatae-Sphagnetum zespół turzycy gwiazdkowatej
Carietum nigrae (subalpinum) (wraz z podzespołem z Juncus filiformis =
Junco filiformis-Sphagnetum recurvi) zespół turzycy pospolitej
Bibliografia
CHYTRÝ M., KUČERA I, KOČI M. (red.) 2001. Katalog bio-topučeske republiky. Agentura Ochrany přirody a krajiny ČR, ss. 304.
DIERSSEN K. 1982. Die wichtigsten Pflanzengesellschaften der Moore in NW-Europas. Conservatoire et Jardin botaniques, Geneve, ss. 382.
DIERSSEN K. 1978. Some aspects of the classification of oligotrophic and mesotrophic mire communities in Europe. Colloques phytosociol. VII, Sols tourbeux, Lille: 399-423.
DIERSSEN K., DIERSSEN B. 1985. Suggestions for a common approach in phytosociology for Scandynavian and European mire ecologist. Aquilo Ser. Bot. 21: 33-44.
HEATHWAITE A. L, EGGELSMANN R., GÖTTLICH K. 1993. Eco-hydrology, Mire Drainage and Mire Conservation. W: GÖTTLICH K. HEATHWAITE A. L. (red.). Mires. Process, Exploitation and Conservation. John Wiley & Sons, Chichester - New York, s: 41 7-476.
HEATHWAITE A. L, GÖTTLICH K., BURMEISTER E. -G., KAULE G., GROSPIETSCH T. 1993. Mires: Definitions and Forms. W: GÖTTLICH K., HEATHWAITE A. L. (red.). Mires. Process, Exploitation and Conservation. John Wiley. & Sons, Chichester - New York, s: 1-75.
JASNOWSKI M. 1962. Budowa i roślinność torfowisk Pomorza Szczecińskiego. Soc. Sc. Stetinensis, 10. ss. 340.
JASNOWSKI M. 1975. Torfowiska i tereny bagienne w Polsce. W: KAC N. J. Bagna kuli ziemskiej. PWN, Warszawa, ss. 475.
JASNOWSKI M., JASNOWSKA J., MARKOWSKI S. 1968. Ginące torfowiska wysokie i przejściowe w pasie nadbałtyckim Polski. Ochr. Przyr. 33: 69-124.
MAREK S., PAŁCZYŃSKI A. 1962. Torfowiska wysokie w Bieszczadach Zachodnich. Zesz. Problem. Post. Nauk Roln,. 34: 255-299.
MATULA J., WOJYTUŃ B., TOMASZEWSKA K., ŻOŁNIERZ L. 1997. Torfowiska polskiej części Karkonoszy i Gór Izerskich. Annales Silesiae 27: 123-140.
MATUSZKIEWICZ W. 2001. Przewodnik do oznaczania zbiorowisk roślinnych Polski. Wyd. Nauk. PWN, ss. 537.
MICHAŁ I. , PETRIČEK V. (red.) 1999. Peče o chranena územi II. Nelesni společenstva. Agentura ochrany přirody a krajiny ČR, ss. 278
OBIDOWICZ A. 1975. Entstehung und Alter einiger Moore im nordlichen Teil der Hohen Tatra - Geneza i wiek kilku torfowisk po północnej stronie Tatr Wysokich. Fragm. Flor. Geobot. ,21,3:289-323.
OBIDOWICZ A. 1985. Torfowiska górskie w Europie. Kosmos 187/2:299-310.
OBIDOWICZ A. 1990. Eine Pollenanalytische und Moorkundli-che Studie zur Vegetationsgeschichte des Podhale - Gebietes (West-Karpaten). Acta Paleobotanice 30, 1-2: 147-219.
PFANDENHAUER J., Schneekloth H., Schneider R., Schneider S. 1993. Mire Distribution. W: Gottlich K., Heathwaite A. L. (red.). Mires. Process, Exploitation and Conservation. John Wiley & Sons, Chichester - New York, s: 77-121.
POTOCKA J. 2000. Stan zachowania oraz geomorfologiczne i hydrologiczne uwarunkowania rozmieszczenia torfowisk w Górach Izerskich. Przyr. Sudetów Zach., 3: 35-44.
RYBNIČEK K., BALATOVA-TULÁČKOVA E. & NEUHÄUSL R. 1984. Přehled rostlinnych společenstev rašelinist’ a mokřadnich luk Československa. Studie ČSAV 8: 5-123.
SEIBERT R 1992. Klasse: YACCINIO - PICEETEA. W: Oberdorfer E. (red.), Suddeutsche Pflanzengesellschaften, T IV, G. Fischer Verl. Jena-Stuttgart-New York, s: 53-80.
TOŁPA S. 1949. Torfowiska Karkonoszy i Gór Izerskich. Roczn. Nauk Roln,. 52, 5-73.
Maria Herbichowa, Joanna Potocka